Evolution & Systematics
Fossil History
The currently oldest known molluscs are found to be small, coiled organisms called Monoplacophora that underwent 180 degrees of torsion of the visceral mass, a characteristic of all gastropods (Ruppert et al. 2004). These organisms were considered to be limpet like gastropods with the presence of a radula and muscular foot and were found in the earliest part of the Cambrian Period at about 545-528 million years ago (mya) with the first cephalopods following at about 495 mya (Bunje 2014). Gastropods and bivalves became more common at the Palaeozoic, but did not obtain dominance of the shallow region of marine environments until the Mesozoic Era (251-65 mya) when they began to greatly increase in population and diversity. Alternative, the cephalopod specie nautiloid, became dominant and gave rise to the current day pearly nautilus (VIC Musuem, 2009). Prior to the nautilus, evolved the ammonoids in the Mesozoic Era. The ammonoids evenutally began to develop an extreme diversity of shell formations and ultimately decreased in abundance to become extinct by the end of the Cretaceous period. Gastropods and bivalves became the most abundant with the greatest biodiversity in the Cainozoic Era less than 65 mya. This is exemplified with the bivalves and gastropods being the most common rock fossilized organisms found in this era that are recognized as similar to present day forms (VIC Museum, 2009). Despite little documentation, it has also been discovered that although the P. ebeninus organisms are currently endemic to Australia, the shells have been found in Tasmanian waters as a sub-fossils beached on the northwestern coast (Grove, 2011).
Gastropod Systematics
Although gastropods are one of the largest and most diverse classes of molluscs, their phylogeny that was considered to be relatively stable is now being reconsidered as their prior configuration did not accurately reflect their evolution (Ruppert et al. 2004). This reconsideration includes the classification of subclasses of Prosobranchia, Opithobranchia, and Pulmonata that are non found to be either paraphyletic or polyphyletic rendering their classifications as unacceptable.
Phylogeny of Potamididae
The evolution of the genus Pyrazus, is highly undocumented. A recent study by Strong (2011) conducted a study on the phylogenetic relationships within the Cerithioidea. This study was used to determine the basal members of the phylogenetic tree (Strong et al. 2011). This analysis in the study determined that Potamididae is the basal member with Paludomidae and Thiaridae as sister taxa with exception of the morphology tree based on the instabilities in the group found in Modulus, Finella, and Scaliola based on the poorly known nature of these particular marine taxa (Fig. 1). As shown in Figure 1, this study classifies P. ebeninus as part of the Batillaridae family, whereas it is classified by all other resources collected as a genus of the Potamididae family. For the purpose of the evolution of this group based on morphological and phylogentic systematics, I will assume that the study completed by Strong (2011) is incorrect on the placement of P. ebeninus and will use the evolution of Potamididae due to lack of supporting resources found on the discrepancy between the two families (Strong et al. 2011). Branching order of family level taxa was found in this study to be most unstable in the group of families of: Batillariidae, Cerithiidae Dialidae, Diastomatidae, Litiopidae, Planazidae, Siliquariidae, Turritellidae. Comparable topologies were also found within the separate group consisting of Modulidae, Paludomidae, Potamididae, Scaliolidae, and Thiaridae.
Both groups studied by Strong (2011) are paraphyletic grades with the exception of Scaliolidae. It is considered that Scaliolidae is the most basal cerithioidean offshoot and was found to be a basal member of latter group in the separate analyses taken (Strong et al. 2011). Thiaridae was found to be basal to a clade containing all remaining Cerithioideans instead of being derived in Group 2 (Fig. 1). It was also discovered that the parasperm of Potamididae is an amophory in the phylogeny tree and that Thiaridae and Paludomidae are not supported as sister taxa shown by their morphological data taken throughout the study. However, it was demonstrated that this finding was supported in all other analyses with exception of the analysis of 28S data.
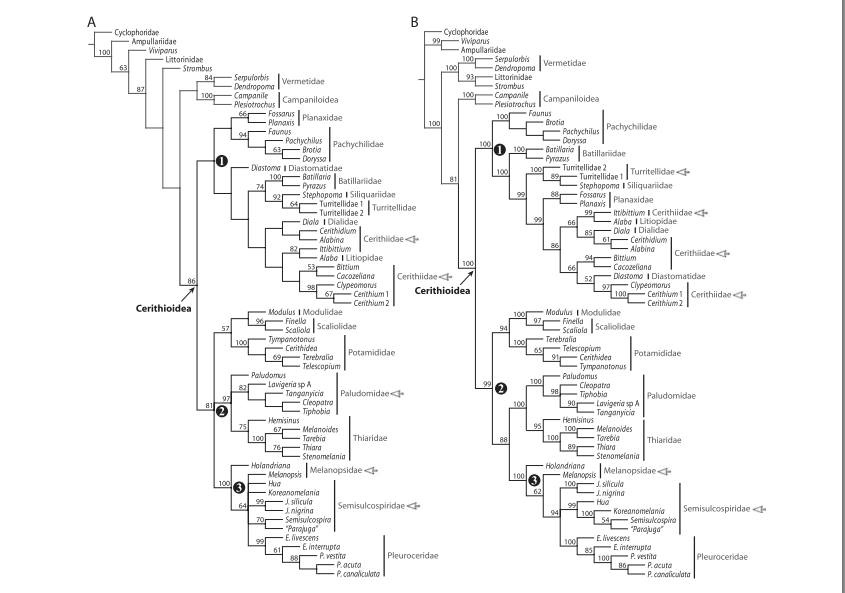
Figure 1: Phylogenetic tree configurations for comparisons of basal morphologies and phylogenies of Group 1 and Group 2 in the study taken by Strong (2011).
|